Bactériophage et phagothérapie
Un bactériophage est un virus n’infectant que des bactéries. En grec, phageton signifie nourriture/consommation. Très utilisés dans la recherche en génétique moléculaire. Les bactériophages sont présents dans l’ensemble de la biosphère. Ils sont présents en quantité plus importante dans les excréments, le sol et les eaux d’égout.
Histoire:
La découverte de l’activité des bactériophages se fait en 1915 par Frederick W. Twort (à Londres) qui remarque que des colonies de microcoques (virus de la vaccine (sorte de variole)) prennent parfois un aspect vitreux, en 24 h à 37 °C dû à une destruction des cellules bactériennes, et que cette caractéristique est transmissible à des colonies normales par simple contact.

En 1917, Félix d’Hérelle fait la même observation dans des selles de malades atteints de dysenterie bacillaire (maladie du côlon), isole les premiers phages, et développe les premières applications thérapeutiques. Le support de l’information génétique (génome) des bactériophages peut être un ADN ou un ARN. Si Twort a découvert les bactériophages, c’est d’Hérelle qui a découvert la phagothérapie.

Dans les années 1940-1960, les travaux effectués sur les bactériophages ont permis de faire de nombreuses avancées dans le domaine de la biologie moléculaire (avancées portées par Max Delbrück, dans le cadre du « groupe phage ») et ont notamment permis de découvrir que les acides nucléiques constituaient le support de l’information génétique (expérience de Hershey-Chase en 1952).

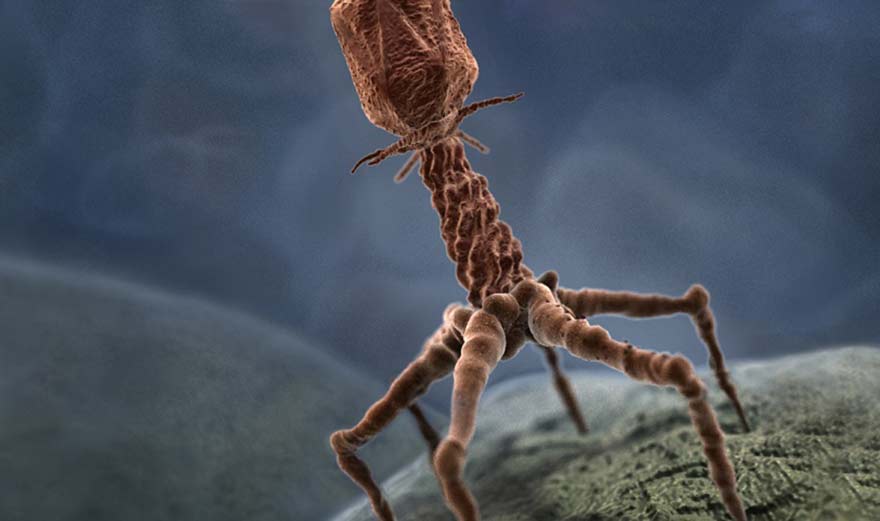
morphologie et structure du bactériophage:
Le virion ne procède qu’un seul type d’acide nucléique : soit de l’ ARN., soit de l’ ADN.
1) Le virion se produit à partir de son seul acide nucléique alors que les autres êtres se reproduisent à partir de la somme de leurs constituants.
2) Le virion est incapable de croître et de subir des divisions binaires.
3) Le virion n’a aucune information génétique concernant les enzymes du métabolisme intermédiaire (susceptibles de produire de l’énergie).
4) La multiplication des virions implique l’utilisation des structures de la cellule hôte, et spécialement de ses ribosomes.
Classification des bactériophages:
Quatre critères essentiels sont retenus:
– La nature des matériels génétiques (ADN ou ARN) ;
– le type de symétrie suivant lequel le virus est construit (hélicoïdale, icosaèdrale, combiné) ;
– le caractère nu ou enveloppé de la nucléo capside.
– les données quantitatives concernant le virion a symétrie icosaèdrale, longueur et épaisseur des nucléo capside pour les virus a symétrie hélicoïdale.
Il y 4 types particulièrement étudiés. La plupart infectent E. coli.
T2:
Ces phages sont à l’origine de beaucoup de manipulations de Biologie moléculaire, les T2 en particulier. (début du 20ème siècle:) A partir de l’étude du T2, HERSHEY et CHASE ont pu mettre en évidence que l’ADN était le support de l’information génétique. D’autre part, VOKIN et ASTRACHAN ont mis en évidence le rôle de l’ARNm et sa fonction biologique.
T7:
Les T7 sont utilisés pour des vecteurs de clonage comme promoteur fort.
Les T pairs comme les T2,4,6 appartiennent à la famille des Myoviridae. A la place de la Cytosine il y a de l’HydroxyMéthylCytosine dans leur génome. L’intérêt est que cela leur permet de résister aux enzymes de restriction lors de l’infection.
Les T impairs appartiennent à la famille des Podoviridae.
Tous les phages de la série T possèdent un ADN linéaire bicaténaire et possèdent une structure avec « tête et queue ».
Ils présentent deux cycles de multiplication, contrairement aux phages T.
-> un cycle lytique, comme les phage T,
-> un cycle lysogène: le phage s’intègre dans le génome bactérien au lieu de rester virion.
Le plus étudié est le phage « lambda » notamment pour les clonage.
Autres: phage P1, P22, P5 et phage Mu.
Ce sont des phages de petite taille avec deux sortes de nucléocapside.
-> nucléocapside icosaédrique (comme X174, S13). L’étude de ces phages a été à l’origine de la découverte des gènes superposés.
-> nucléocapside filamenteuse (comme phage fd, phage M13). Ces phages sont très utilisés car ADN simple brin (donc facilement séquençable).
Ils appartiennent à la famille des Leviridae comme le phage Qß, Ms2. Ils servent de modèles pour la traduction de l’ARN.
Les phages présentent différents types de génomes: ADN double-brins, ADN simple-brin, ARN; et différents types de structures de nucléocapside: filamenteuse, isocaïdrique ou comme les T2.
Depuis les surprenantes images de bactériophages « en forme de têtards » observées au microscope électronique par les premiers expérimentateurs (Ruska, en 1941, Lurai Delburck et Anderson en 1941), le développement considérable des recherches sur la structure des bactériophages a conduit à la description de nombreuses variétés morphologiques et à l’élaboration d’une véritable anatomie ultrastructurale de ces virus.
Actuellement en peut réunir les bactériophages en trois grands groupes morphologiques :
– cubique.
– Filamenteux.
– Mixte.

Le phage lambda (λ ) est un virus à structure mixte. Le diamètre de la tête et de 100 mm. Les virions sont constitués par 12 protéines structurales localisées dans l’enveloppe (P9, P10 et P13), Peplomers (P3 et P6), nucléocapside (P8 et P5), complexe polymérase (P1, P2, P4 et 97) et une protéine non structurale P12. Les protéines constituent environ 70% du poids de la particule.
Deux Types de Cycles.
Ici le phage se multiplie au dépend de la bactérie et généralement la détruit. Ces sortes de phages sont dits « virulents ».
Ici le génome du phage est injecté dans la bactérie, mais ne s’y réplique pas. Par contre il va s’intégrer dans le génome bactérien. La multiplication du phage se fait grâce à la réplication du chromosome bactérien. Le phage s’intègre à un seul endroit du génome bactérien.
Quand le phage est intégré, il y a un phénomène d’immunité qui préserve la bactérie d’une surinfection du même phage.
Un phage intégrer = prophage et seuls les phages tempérés ont la capacité d’avoir un cyle lysogène (ils possèdent aussi un cycle lytique).
Le phage se fixe sur la bactérie. C’est une phase fondamentale car il y a une reconnaissance spécifique entre le récepteur et le phage (sinon pas de fixation). Les conditions d’absorption dépendent, parfois des cofacteurs libérés par la bactérie qui va être parasitée.
Ex: T4 se fixe bien si la bactérie libère Tyr.
« lambda » se fixe bien si la bactérie libère Mg2+.
Le récepteur, au niveau de la bactérie, varie suivant le phage.
T2 = porine; « lambda » = porine lam B.
On retrouve ainsi des pilus sexuels (F) pour M13. Certains utilisent le LPS comme site de fixation.
Pour le phage T2, la fixation est effectuée par les filament codaux et la plaque terminale. Elles devient rapidement irréversible. C’est ici que les enzymes de la plaque codale s’activent et hydrolysent les enveloppes bactériennes pour pouvoir injecter l’ADN virale.
Elle est réalisée par contraction de la gaine externe qui favorise la pénétration du cylindre central.
Après injection, le virion cesse d’exister, ne persiste que le génome dans la bactérie. Il n’y a plus de particule virale (environ 12 min). De nombreux évenements se déroulent alors: la synthèse de l’ADN bactérien est stoppé par le phage, une DNAse phagique va dégrader le génome bactérien (DNAse phagique pendant 2 à 3 min après l’injection , issue des gènes précoces). Il y a aussi production d’ARNm et production de protéines constituant la nucléocapside et, parallèlement l’ADN se mulitiplie. Cet ADN est formé sous forme de « concatemères » (= cercle roulant) c’est-à dire plusieurs séquences d’ADN collées à la suite. Ici accumulation des différents composants du phage: ADN phagique, nucléocapside => finde la phase d’éclipse.
Entre 12 à 15 min. L’ADN produit en concatemère est coupé en fragments de taille normale avec parfois des sources d’erreurs (101 à 112% taille totale du génome) donc fragments plus longs ou plus courts. Ce qui est à l’origine d’une importante mutation des phages. La nucléocapside se structure, il y a alors apparition de la particule virale = virions dans la bactérie.
Quand la bactérie accumule entre 100 à 200 virions, il y a activation d’une endolysine phagique qui entraîne l’éclatement de la bactérie et la libération des phages.
La strucutre est « tête et queue » mais ici le phage ne possède pas de filaments codaux. Ici, l’expression du phage est réprimée. Pour sortir de cette phase lysogène il faut que la bactérie hôte subisse « un stress » (rayons X, UV, agent mutagène,…) alors le phage devient productif -> cycle lytique.
Il s’agit d’un phage qui infecte E. Coli K12. Il possède une génome double-brin à extrémités cohésives (donc circulaire).
Lorsque le génome du phage est infecté dans la bactérie il se circularise.
Taille » 49Kpb.
- 1. — Gènes impliqués dans la régulation de lysogénie.
2. — Gènes impliqués dans la recombinaison, c’est-à dire l’étape d’intégration (aH: séquence complémentaire qui permet l’intégration).
3. — Gènes impliqués dans la synthèse de m’ADN (lytique).
4. — Gènes impliqués dans le phénomène de lyse de la bactérie (lytique).
Tout le reste sont des gènes qui codent pour la nucléocapside, des gènes structuraux.
Il débute comme le phage T2.
Résistance à la bactérie = Lam B.
Quand le génome est injecté, il se circuralise et il débute sa transcription. Protéine Cro et N dès le début.
Cro = régule les cyles lysogène / lytique.
N = permet la synthèse d’ARNm de taille variable « aterminateur ».
Puis CII, CIII et Q –> switch –> soit lysogène soit lytique.
Dans la phase lytique [Q] augmente ce qui entraîne l’augementation du régulateur Pro: on bloque alors les protéines qui activent la lysogénie (rétocontrôle +). Le phage produit alors des ARNm longs et donc fabrique sa tête et sa queue.
Comme le phage T2, l’ADN est répliqué sous forme de contatemère puis est réduis par une DNAse virale qui génère les extrémités cohésives (ex: EcoR1).
Pour qu’il soit induit il faut réprimer les gènes tardifs, qui codent pour le tête et la queue et activer les gènes de recombinaison. La protéine régulatrice est CI. Par un système de régulation génique, CI s’exprime et augmente ce qui inhibe les gènes tardifs. Il y a alors activation du cycle lysogène ou le génome viral s’intègre au génome bactérien. La sortie de cette phase lysogène (phase naturelle du phage) se fait par REC A (protéine bactérienne) qui est produit en cas de stress (RECA hydrolyse CI ce qui induit le cycle lytique).
-
Diagnostic et Taxonomie:
Principe de la lysotypie: on s’en sert au niveau médical, au niveau industriel pour identifier les couches de levain. La recherche du phage est un indicateur de Contamination Fécale (eau, aliments,…).
- M12 possède un ADN monobrin donc utile pour le séquençage. ll est utilisé comme vecteur de clonage (phagemide) et comme un système de traduction in vitro (production d’ARNm).
- la phagothérapie moyen pour lutter contre des infections bactériennes comme la maladie de Lyme?
La Borrelia responsable de cette maladie se soigne par antibiotiques, mais les traitements sont lourds et ne l’éliminent pas toujours. De plus, faisant partie des bactéries les plus rapides et les plus mobiles que l’on connaisse, on la sait capable de se loger dans tous les tissus du corps humain, se rendant comme invisible et, de ce fait, inatteignable par les antibiotiques. En effet, pendant toute la durée du traitement, elle peut se localiser sous forme de kyste spongieux, attirant les globules blancs qui finissent par s’agglutiner et ainsi la dissimuler. La bactérie s’en sort saine et sauve en laissant des lésions créées par l’amas de globules blancs.
En 1982, Hayes, Burgdorfer et Barbour ont enregistré et observé l’attaque d’un phage contre Borrelia burgdorferi in vivo et ont pris des photographies pour enregistrer l’événement. Les images capturées sont celles d’un bactériophage de type B3, décrit par les chercheurs comme ayant une « tête allongée de 40 à 50 nm et une queue de 50 à 70 nm de long. Il semble dépourvu de collets ou de structure en cerf-volant ».
En 2001, Eggers et al ont publié leur découverte d’un phage de Borrelia burgdorferi (Bb) nommé phiBB-1 (également appelé φBB-1). Ce n’est pas le meilleur candidat pour une utilisation dans le traitement par les bactériophages, car il s’agit d’un phage lysogène tempéré.
Les phages lysogènes restent inactifs en tant que virus lorsqu’ils sont des prophages et ne se répliquent qu’avec le génome de l’hôte, à moins d’être mobilisés. En revanche, les phages virulents, ayant été répliqués et assemblés en virions complets, provoquent une lyse rapide et la mort de la cellule bactérienne, avec une libération de 10 à 100 virions par phage. Ces virions trouvent alors plus de proies et disparaissent lorsqu’elles ne peuvent plus trouver de bactéries.
Malheureusement, à ce jour, ni les scientifiques, ni aucun centre de recherche n’ont travaillé sur le traitement des phages contre les souches de Borrelia.
L’un des obstacles majeur dans la recherche et la mise sur le marché de produits de phagothérapie est l’hyper spécificité des bactériophages à chaque souche de bactérie et donc peu adaptés au développement industriel, et par ailleurs difficilement brevetables.
Article en rapport avec ce sujet
https://www.francetvinfo.fr/sante/decouverte-scientifique/infection-bacterienne-une-jeune-fille-sauvee-grace-a-des-phages_3442843.html
Demande de devis
Comparez des devis pour votre projet écologique
Décrivez votre besoin et recevez des propositions adaptées via Leadrs.